
Visas „iLive“ turinys yra peržiūrėtas medicinoje arba tikrinamas, kad būtų užtikrintas kuo didesnis faktinis tikslumas.
Mes turime griežtas įsigijimo gaires ir susiejamos tik su geros reputacijos žiniasklaidos svetainėmis, akademinių tyrimų institucijomis ir, jei įmanoma, medicininiu požiūriu peržiūrimais tyrimais. Atkreipkite dėmesį, kad skliausteliuose ([1], [2] ir tt) esantys numeriai yra paspaudžiami nuorodos į šias studijas.
Jei manote, kad bet koks mūsų turinys yra netikslus, pasenęs arba kitaip abejotinas, pasirinkite jį ir paspauskite Ctrl + Enter.
Kiaušidės
Medicinos ekspertas

Paskutinį kartą peržiūrėta: 04.07.2025

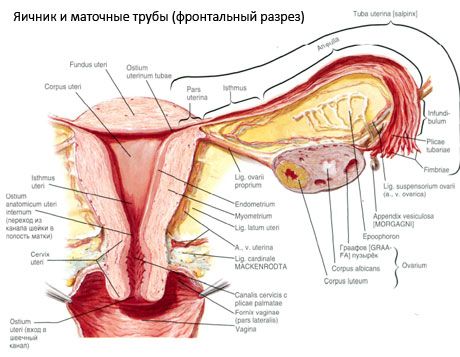
Kiaušidė (ovarium; graikiškai oophoron) yra porinis organas, moters lytinė liauka, esanti dubens ertmėje už plačiojo gimdos raiščio. Kiaušidėse vystosi ir bręsta moters lytinės ląstelės (kiaušialąstės), susidaro moteriški lytiniai hormonai, kurie patenka į kraują ir limfą. Kiaušidė yra ovalo formos, šiek tiek suplokštėjusi priekine ir užpakaline kryptimi. Kiaušidės spalva rausva. Pagimdžiusios moters kiaušidės paviršiuje matomi įdubimai ir randai – ovuliacijos ir geltonkūnio transformacijos pėdsakai. Kiaušidė sveria 5–8 g. Kiaušidės matmenys: ilgis 2,5–5,5 cm, plotis 1,5–3,0 cm, storis – iki 2 cm. Kiaušidė turi du laisvus paviršius: medialinį paviršių (facies medialis), nukreiptą į dubens ertmę, iš dalies uždengtą kiaušintakiu, ir šoninį paviršių (facies lateralis), esantį greta dubens šoninės sienelės, iki šiek tiek ryškaus įdubimo – kiaušidės duobės. Ši duobė yra kampe tarp išorinių klubakaulių kraujagyslių, viršuje padengtų pilvaplėve, ir gimdos bei obturatoriaus arterijų apačioje. Už kiaušidės atitinkamos pusės šlapimtakis eina retroperitoniniu būdu iš viršaus į apačią.
Kiaušidės paviršiai pereina į išgaubtą laisvą (užpakalinį) kraštą (margo liber), priekyje – į žarnų pasaito kraštą (margo mesovaricus), trumpu pilvaplėvės (kiaušidės žarnų pasaito) raukšle pritvirtintą prie gimdos plačiojo raiščio užpakalinio lapelio. Ant šio priekinio organo krašto yra griovelių išraižyta įduba – kiaušidės „hilum“ (hilum ovarii), per kurią į kiaušidę patenka arterija ir nervai, o išeina venos ir limfagyslės. Kiaušidė taip pat turi du galus: viršutinį apvalų vamzdinį galą (extremitas tubaria), nukreiptą į kiaušintakį, ir apatinį gimdos galą (extremitas utenna), sujungtą su gimda kiaušidės raiščiu (lig. ovarii proprium). Šis maždaug 6 mm storio apvalios virvelės formos raištis eina nuo kiaušidės gimdos galo iki gimdos šoninio kampo, esančio tarp dviejų gimdos plačiojo raiščio lapelių. Kiaušidės raiščių aparatui taip pat priklauso kiaušidės raiščio suspensija (lig.suspensorium ovarii) – pilvaplėvės raukšlė, einanti nuo mažojo dubens sienelės iki kiaušidės, kurios viduje yra kiaušidės kraujagyslės ir skaidulinių skaidulų pluoštai. Kiaušidę fiksuoja trumpas žarnų pasaitas (mezovariumas) – pilvaplėvės dubenėlis, einantis nuo gimdos plačiojo raiščio užpakalinio lapelio iki kiaušidės žarnų pasaito krašto. Pačių kiaušidžių pilvaplėvė nedengia. Didžiausias kiaušintakio kraštas yra pritvirtintas prie kiaušidės vamzdinio galo. Kiaušidės topografija priklauso nuo gimdos padėties, jos dydžio (nėštumo metu). Kiaušidės yra labai judrūs mažojo dubens ertmės organai.
 [
[ Kiaušidžių indai ir nervai
Kiaušides krauju aprūpina aa. ir vv. ovaricae et uterinae. Abi kiaušidžių arterijos (aa. ovaricae dextra et sinistra) prasideda nuo priekinio aortos paviršiaus, tiesiai po inkstų arterijomis; dešinioji paprastai prasideda nuo aortos, o kairioji – nuo inkstų arterijos. Kiekviena kiaušidžių arterija, eidama žemyn ir į šonus išilgai didžiojo juosmens raumens priekinio paviršiaus, kerta priekyje esantį šlapimtakį (atšakodama jam), išorinius klubakaulius, ribą ir įeina į dubens ertmę, čia būdama kiaušidės pakabos raištyje. Medialine kryptimi kiaušidžių arterija praeina tarp plačiojo gimdos raiščio lapelių po kiaušintakiu, atšakodama jam, o tada į kiaušidės žarnas; ji įeina į kiaušidės žarną.
Kiaušidės arterijos šakos plačiai anastomozuoja su gimdos arterijos kiaušidės šakomis. Veninis nutekėjimas iš kiaušidžių pirmiausia atliekamas į kiaušidžių venų rezginį, esantį kiaušidės kauburėlio srityje. Iš čia kraujas teka dviem kryptimis: per gimdos ir kiaušidės venas. Dešinė kiaušidės vena turi vožtuvus ir įteka į apatinę tuščiąją veną. Kairioji kiaušidės vena įteka į kairę inkstų veną, nors ir neturi vožtuvų.
Limfos drenažas iš kiaušidžių vyksta per limfagysles, ypač gausiai organų vartų srityje, kur išsiskiria subovarinis limfinis rezginys. Tada limfa išleidžiama kiaušidžių limfagyslėmis į paraaortinius limfmazgius.
Kiaušidžių inervacija
Simpatinė – teikiama postganglioninių skaidulų iš celiakijos (saulės), viršutinio mezenterinio ir hipogastrinio rezginių; parasimpatinė – per visceralinius sakralinius nervus.
Kiaušidės struktūra
Kiaušidės paviršių dengia vienasluoksnis gemalinis epitelis. Po juo slypi tankus jungiamojo audinio baltyminis apvalkalas (tunica albuginea). Kiaušidės jungiamasis audinys sudaro jos stromą (stroma ovarii), kurioje gausu elastingų skaidulų. Kiaušidės medžiaga, jos parenchima, yra padalinta į išorinį ir vidinį sluoksnius. Vidinis sluoksnis, esantis kiaušidės centre, arčiau jos vartų, vadinamas šerdimi (medulla ovarii). Šiame sluoksnyje, puriame jungiamajame audinyje, yra daugybė kraujagyslių ir limfagyslių bei nervų. Išorinis kiaušidės sluoksnis – žievė (cortex ovarii) – yra tankesnė. Joje daug jungiamojo audinio, kuriame išsidėstę bręstantys pirminiai kiaušidžių folikulai (folliculi ovarici primarii), antriniai (pūsliniai) folikulai (folliculi ovarici secundarii, s.vesiculosi), taip pat subrendę folikulai, Grafo folikulai (folliculi ovarici maturis), taip pat geltonieji ir atretiniai kūneliai.
Kiekviename folikule yra moteriškas reprodukcinis kiaušinėlis arba oocitas (oocitas). Kiaušialąstė yra iki 150 µm skersmens, apvali, turi branduolį, daug citoplazmos, kurioje, be ląstelės organelių, yra baltymų ir lipidų intarpų (trynio), glikogeno, būtinų kiaušialąstės mitybai. Kiaušialąstė paprastai sunaudoja savo maistinių medžiagų atsargas per 12–24 valandas po ovuliacijos. Jei apvaisinimas neįvyksta, kiaušialąstė žūsta.
Žmogaus kiaušinėlis turi dvi membranas, dengiančias jį. Viduje yra citolema – kiaušinėlio citoplazminė membrana. Už citolemos yra vadinamųjų folikulinių ląstelių sluoksnis, kuris apsaugo kiaušinėlį ir atlieka hormonų formavimo funkciją – išskiria estrogenus.
Gimdos, kiaušintakių ir kiaušidžių fiziologinę padėtį užtikrina pakabinimo, fiksavimo ir atramos aparatai, jungiantys pilvaplėvę, raiščius ir dubens audinį. Pakabinimo aparatas yra porinis darinys, jį sudaro apvalieji ir platieji gimdos raiščiai, tikri raiščiai ir kiaušidžių pakabinimo raiščiai. Plačiieji gimdos raiščiai, tikri ir pakabinimo raiščiai laiko gimdą vidurinėje padėtyje. Apvalieji raiščiai traukia gimdos dugną į priekį ir suteikia jos fiziologinį pakreipimą.
Fiksavimo (įtvirtinimo) aparatas užtikrina gimdos padėtį mažojo dubens centre ir praktiškai neleidžia jai pasislinkti į šonus, pirmyn ir atgal. Tačiau kadangi raiščių aparatas nuo gimdos atsitraukia apatinėje dalyje, gimda gali pakrypti įvairiomis kryptimis. Fiksavimo aparatą sudaro raiščiai, esantys laisvuosiuose dubens audiniuose ir besitęsiantys nuo apatinės gimdos dalies iki šoninės, priekinės ir užpakalinės dubens sienelių: kryžkaulio-magijos, kardininiai, gimdos-vezikiniai ir vezikopubiniai raiščiai.
Be mezovariumo, išskiriami šie kiaušidžių raiščiai:
- Kiaušidės pakabos raištis, anksčiau vadintas infundibulopviliniu raiščiu. Tai pilvaplėvės raukšlė su per ją einančiomis kiaušidės kraujagyslėmis (a. et v. ovarica), limfagyslėmis ir nervais, ištempta tarp dubens šoninės sienelės, juosmens fascijos (bendrosios klubakaulio arterijos dalijimosi į išorinę ir vidinę srityje) ir viršutinio (kiaušintakio) kiaušidės galo;
- Tinkamas kiaušidės raištis eina tarp plačiojo gimdos raiščio lapelių, arčiau užpakalinio lapelio, ir jungia apatinį kiaušidės galą su gimdos šoniniu kraštu. Tinkamas kiaušidės raištis yra prisitvirtinęs prie gimdos tarp kiaušintakio pradžios ir apvaliojo raiščio, už ir virš pastarojo. Raiščio storiu praeina rr. ovarii, kurie yra gimdos arterijos galinės šakos;
- Apendikulinis-kiaušidžių raištis. Raištis tęsiasi nuo apendikso žarnų iki dešiniosios kiaušidės arba plačiojo gimdos raiščio pilvaplėvės raukšlės pavidalu. Raištis yra nepastovus ir stebimas 1/2–1/3 moterų.
Atraminį aparatą vaizduoja dubens dugno raumenys ir fascija, suskirstyti į apatinį, vidurinį ir viršutinį (vidinį) sluoksnius.
Galingiausias yra viršutinis (vidinis) raumenų sluoksnis, kurį sudaro porinis raumuo, keliantis išangę. Jį sudaro raumenų pluoštai, kurie išsidėstę nuo uodegikaulio iki dubens kaulų trimis kryptimis (pubokokeinis raumuo, klubiniskokeinis raumuo ir ischiokokeinis raumuo). Šis raumenų sluoksnis dar vadinamas dubens diafragma.
Vidurinis raumenų sluoksnis yra tarp simfizės, gaktos ir sėdynkaulių. Vidurinis raumenų sluoksnis – urogenitalinė diafragma – užima priekinę dubens išėjimo pusę, per kurią praeina šlaplė ir makštis. Priekinėje dalyje tarp jos lakštų yra raumenų ryšuliai, sudarantys išorinį šlaplės sfinkterį, užpakalinėje dalyje – raumenų ryšuliai, einantys skersine kryptimi – gilusis skersinis tarpvietės raumuo.
Apatinis (išorinis) dubens dugno raumenų sluoksnis susideda iš paviršinių raumenų, kurių forma primena skaičių 8. Tai apima bulbokaverninį, ischioakaverninį, išorinį analinį sfinkterį ir paviršinį skersinį tarpvietės raumenį.
Kiaušidžių ontogenezė
Folikulų augimo ir atrezijos procesas prasideda 20 nėštumo savaitę, o iki gimdymo mergaitės kiaušidėse lieka iki 2 milijonų oocitų. Iki menarchės jų skaičius sumažėja iki 300 tūkstančių. Per visą reprodukcinio gyvenimo laikotarpį subręsta ir ovuliuoja ne daugiau kaip 500 folikulų. Pradinis folikulų augimas nepriklauso nuo FSH stimuliacijos, yra ribotas, o atrezija įvyksta greitai. Manoma, kad vietoj steroidinių hormonų pagrindinis pirminių folikulų augimo ir atrezijos reguliatorius yra vietiniai autokriniai/parakriniai peptidai. Manoma, kad folikulų augimo ir atrezijos proceso nepertraukia jokie fiziologiniai procesai. Šis procesas tęsiasi visais amžiais, įskaitant intrauterininį laikotarpį ir menopauzę, ir jį pertraukia nėštumas, ovuliacija ir anovuliacija. Mechanizmas, kuris sukelia folikulų augimą ir jų skaičių kiekviename konkrečiame cikle, vis dar nėra aiškus.
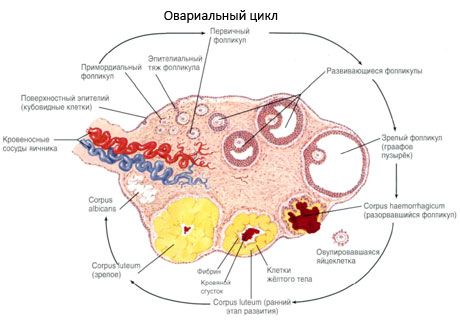
Folikulo vystymosi metu vyksta keli vystymosi etapai. Pirmykštės gemalo ląstelės atsiranda iš trynio maišelio endodermos, alantoiso, ir 5–6 nėštumo savaitę migruoja į embriono lytinių organų sritį. Dėl greito mitozinio dalijimosi, kuris tęsiasi nuo 6–8 iki 16–20 nėštumo savaičių, embriono kiaušidėse susidaro iki 6–7 milijonų oocitų, apsuptų plonu granuliuotų ląstelių sluoksniu.
Preantralinis folikulas – oocitą supa membrana (Zona pellucida). Oocitą supančios granuliuotos ląstelės pradeda daugintis, jų augimas priklauso nuo gonadotropinų ir koreliuoja su estrogenų kiekiu. Granuliuotos ląstelės yra FSH taikinys. Preantralinio folikulo stadijoje granuliuotos ląstelės geba sintetinti trijų klasių steroidus: daugiausia indukuoja aromatazės, pagrindinio fermento, kuris androgenus paverčia estradioliu, aktyvumą. Manoma, kad estradiolis geba padidinti savo receptorių skaičių, darydamas tiesioginį mitogeninį poveikį granuliuotoms ląstelėms, nepriklausomą nuo FSH. Jis laikomas parakrininiu faktoriumi, kuris sustiprina FSH poveikį, įskaitant aromatizacijos procesų aktyvavimą.
FSH receptoriai atsiranda granuliuotų ląstelių membranose, kai tik prasideda folikulų augimas. FSH sumažėjimas arba padidėjimas lemia jo receptorių skaičiaus pokytį. Šį FSH poveikį moduliuoja augimo faktoriai. FSH veikia per G baltymą, adenilato ciklazės sistemą, nors steroidogenezę folikule daugiausia reguliuoja FSH, šiame procese dalyvauja daug veiksnių: jonų kanalai, tirozino kinazės receptoriai, antrųjų pasiuntinių fosfolipazės sistema.
Androgenų vaidmuo ankstyvajame folikulų vystymesi yra sudėtingas. Granulinės ląstelės turi androgenų receptorius. Jie yra ne tik FSH sukeltos aromatizacijos estrogenais substratas, bet ir gali sustiprinti aromatizaciją esant mažoms koncentracijoms. Kai padidėja androgenų kiekis, preantralinės granulinės ląstelės pirmiausia renkasi ne aromatizacijos kelią į estrogenus, o paprastesnį androgenų kelią per 5α-reduktazę, kad paverstų juos androgenu, kuris negali būti paverstas estrogenu, taip slopindamos aromatazės aktyvumą. Šis procesas taip pat slopina FSH ir LH receptorių susidarymą, taip sustabdydamas folikulų vystymąsi.
Aromatizacijos proceso metu folikulas, kuriame yra daug androgenų, patiria atrezijos procesus. Folikulo augimas ir vystymasis priklauso nuo jo gebėjimo androgenus paversti estrogenais.
Esant FSH, dominuojanti folikulinio skysčio medžiaga bus estrogenai. Nesant FSH – androgenai. LH folikuliniame skystyje paprastai nėra iki ciklo vidurio. Kai tik padidėja LH lygis, sumažėja granuliuotųjų ląstelių mitozinis aktyvumas, atsiranda degeneracinių pokyčių ir padidėja androgenų lygis folikule. Steroidų lygis folikuliniame skystyje yra didesnis nei plazmoje ir atspindi kiaušidžių ląstelių: granuliuotųjų ir theka ląstelių, funkcinį aktyvumą. Jei vienintelis FSH taikinys yra granuliuotosios ląstelės, tai LH turi daug taikinių – theka ląsteles, stromos ir liuteino ląsteles bei granuliuotasias ląsteles. Tiek granuliuotosios, tiek theka ląstelės gali vykdyti steroidogenezę, tačiau granuliuotosiose ląstelėse vyrauja aromatazės aktyvumas.
Reaguodamos į LH, theca ląstelės gamina androgenus, kuriuos granulosa ląstelės vėliau paverčia estrogenais per FSH sukeltą aromatizaciją.
Folikului augant, thekos ląstelės pradeda ekspresuoti LH receptorių, P450 sec ir 3beta-hidroksisteroidų dehidrogenazės genus, insuliną primenantis augimo faktorius (IGF-1) sinergiškai su LH padidina genų ekspresiją, bet nestimuliuoja steroidogenezės.
Kiaušidžių steroidogenezė visada priklauso nuo LH. Folikului augant, theca ląstelės ekspresuoja fermentą P450c17, kuris iš cholesterolio sudaro androgeną. Granulosa ląstelės šio fermento neturi ir priklauso nuo theca ląstelių, kad jos gamintų estrogenus iš androgenų. Skirtingai nuo steroidogenezės, folikulogenezė priklauso nuo FSH. Folikului augant ir didėjant estrogenų kiekiui, suaktyvėja grįžtamojo ryšio mechanizmas – slopinama FSH gamyba, o tai savo ruožtu lemia folikulo aromatazės aktyvumo sumažėjimą ir galiausiai folikulo atreziją per apoptozę (programuotą ląstelių mirtį).
Grįžtamojo ryšio mechanizmas, veikiantis estrogenus ir FSH, slopina pradėjusių augti folikulų, bet ne dominuojančio folikulo, vystymąsi. Dominuojančiame folikule yra daugiau FSH receptorių, kurie palaiko granuliuotų ląstelių proliferaciją ir androgenų aromatizaciją į estrogenus. Be to, parakrininis ir autokrinis keliai veikia kaip svarbus antralinių folikulų vystymosi koordinatorius.
Autokrinis/parakrinis reguliatorius susideda iš peptidų (inhibino, aktyvino, folistatino), kuriuos sintetina granulosa ląstelės reaguodamos į FSH, ir kurie patenka į folikulinį skystį. Inhibinas mažina FSH sekreciją; aktyvinas stimuliuoja FSH išsiskyrimą iš hipofizės ir sustiprina FSH veikimą kiaušidėje; folistatinas slopina FSH aktyvumą, galbūt prisijungdamas prie aktyvino. Po ovuliacijos ir geltonkūnio išsivystymo inhibiną kontroliuoja LH.
Kiaušidžių ląstelių augimą ir diferenciaciją įtakoja insulino tipo augimo faktoriai (IGE). IGF-1 veikia granuliuotas ląsteles, sukeldamas ciklinio adenozino monofosfato (cAMP), progesterono, oksitocino, proteoglikano ir inhibino kiekio padidėjimą.
IGF-1 veikia theca ląsteles, sukeldamas padidėjusią androgenų gamybą. Theca ląstelės, savo ruožtu, gamina naviko nekrozės faktorių (TNF) ir epidermio augimo faktorių (EGF), kuriuos taip pat reguliuoja FSH.
EGF stimuliuoja granulosa ląstelių proliferaciją. IGF-2 yra pagrindinis folikulinio skysčio augimo faktorius, jame taip pat randami IGF-1, TNF-α, TNF-3 ir EGF.
Kiaušidžių funkcijos parakrininės ir (arba) autokrinės reguliacijos sutrikimas, atrodo, vaidina svarbų vaidmenį ovuliacijos sutrikimams ir policistinių kiaušidžių vystymuisi.
Augant antraliniam folikului, estrogenų kiekis folikuliniame skystyje didėja. Didžiausio padidėjimo metu granuliuotose ląstelėse atsiranda LH receptorių, vyksta granuliuotų ląstelių liuteinizacija ir padidėja progesterono gamyba. Taigi, preovuliaciniu laikotarpiu padidėjusi estrogenų gamyba sukelia LH receptorių atsiradimą, o LH savo ruožtu sukelia granuliuotų ląstelių liuteinizaciją ir progesterono gamybą. Padidėjęs progesteronas sumažina estrogenų kiekį, o tai, matyt, sukelia antrą FSH piką ciklo viduryje.
Manoma, kad ovuliacija įvyksta praėjus 10–12 valandų po LH piko ir 24–36 valandoms po estradiolio piko. Manoma, kad LH stimuliuoja oocito dalijimąsi, granuliuotų ląstelių liuteinizaciją ir progesterono bei prostaglandinų sintezę folikule.
Progesteronas sustiprina proteolitinių fermentų aktyvumą, kurie kartu su prostaglandinu dalyvauja folikulo sienelės plyšime. Progesterono sukeltas FSH pikas leidžia oocitui išeiti iš folikulo, plazminogeną paverčiant proteolitiniu fermentu plazminu, ir suteikia pakankamą LH receptorių skaičių normaliam liuteininės fazės vystymuisi.
Per 3 dienas po ovuliacijos granuliuotos ląstelės padidėja ir jose atsiranda būdingų vakuolių, pripildytų pigmento liuteino. Tekaluteininės ląstelės diferencijuojasi nuo tekos ir stromos ir tampa geltonkūnio dalimi. Kapiliarai, prasiskverbiantys į geltonkūnį, labai greitai vystosi veikiant angiogenezės faktoriams, o pagerėjus vaskuliarizacijai, padidėja progesterono ir estrogenų gamyba. Steroidogenezės aktyvumą ir geltonkūnio gyvavimo trukmę lemia LH lygis. Geltonkūnis nėra homogeniškas ląstelinis darinys. Be 2 tipų geltonkūnio ląstelių, jame yra endotelio ląstelių, makrofagų, fibroblastų ir kt. Didelės geltonkūnio ląstelės gamina peptidus (relaksiną, oksitociną) ir yra aktyvesnės steroidogenezėje, pasižymi didesniu aromatazės aktyvumu ir didesne progesterono sinteze nei mažos ląstelės.
Progesterono pikas pasiekiamas 8 dieną po LH piko. Pažymėtina, kad progesteronas ir estradiolis liuteininėje fazėje epizodiškai išskiriami, koreliuojant su pulsuojančia LH išsiskyrimu. Susiformavus geltonkūniui, inhibino gamybos kontrolė pereina iš FSH į LH. Inhibino kiekis didėja kartu su estradiolio kiekio padidėjimu prieš LH piką ir toliau didėja po LH piko, nors estrogeno kiekis mažėja. Nors inhibiną ir estradiolį išskiria granuliuotos ląstelės, jų kiekį reguliuoja skirtingi keliai. Inhibino kiekio sumažėjimas liuteininės fazės pabaigoje prisideda prie FSH padidėjimo kitam ciklui.
Geltonkūnis mažėja labai greitai – 9–11 dieną po ovuliacijos.
Degeneracijos mechanizmas nėra aiškus ir nėra susijęs su estrogenų liuteolitiniu vaidmeniu ar su receptoriais susijusiu mechanizmu, kuris pastebėtas endometriume. Yra ir kitas geltonkūnio gaminamų estrogenų vaidmens paaiškinimas. Yra žinoma, kad estrogenai yra būtini progesterono receptorių sintezei endometriume. Liuteininės fazės estrogenai tikriausiai yra būtini su progesteronu susijusiems pokyčiams endometriume po ovuliacijos. Nepakankamas progesterono receptorių išsivystymas dėl nepakankamo estrogenų kiekio galimai yra papildomas nevaisingumo ir ankstyvo persileidimo mechanizmas – kita liuteininės fazės trūkumo forma. Manoma, kad geltonkūnio gyvenimo trukmė nusistovi ovuliacijos metu. Ir jis tikrai regresuos, jei dėl nėštumo jo nepalaiko žmogaus chorioninis gonadotropinas. Taigi, geltonkūnio regresija lemia estradiolio, progesterono ir inhibino kiekio sumažėjimą. Inhibino sumažėjimas panaikina jo slopinamąjį poveikį FSH; Estradiolio ir progesterono kiekio sumažėjimas leidžia labai greitai atkurti GnRH sekreciją ir pašalinti grįžtamojo ryšio mechanizmą iš hipofizės. Inhibino ir estradiolio kiekio sumažėjimas kartu su GnRH kiekio padidėjimu sukelia FSH, o ne LH, išsiskyrimą. FSH padidėjimas sukelia folikulų augimą, po kurio seka dominuojančio folikulo atranka, o jei nėštumas neįvyksta, prasideda naujas ciklas. Steroidiniai hormonai vaidina pagrindinį vaidmenį reprodukcinėje biologijoje ir bendrojoje fiziologijoje. Jie lemia žmogaus fenotipą, veikia širdies ir kraujagyslių sistemą, kaulų metabolizmą, odą, bendrą savijautą ir vaidina pagrindinį vaidmenį nėštumo metu. Steroidinių hormonų veikimas atspindi tarpląstelinius ir genetinius mechanizmus, kurie yra būtini norint perduoti tarpląstelinį signalą į ląstelės branduolį, kad sukeltų fiziologinį atsaką.
Estrogenai difuziškai prasiskverbia pro ląstelės membraną ir jungiasi prie receptorių, esančių ląstelės branduolyje. Tada receptorių ir steroidų kompleksas jungiasi prie DNR. Tikslinėse ląstelėse ši sąveika sukelia genų ekspresiją, baltymų sintezę ir specifines ląstelių bei audinių funkcijas.
Использованная литература